
ORF3a
ORF3a is an integral transmembrane protein, localized mostly in the Golgi complex, cytoplasm and cell surface. ORF3a is able to form homotetramers with ion channel properties. Regarding pathogenic effects, ORF3a is linked to inflammatory responses, weak INF responses, innate immunity responses, trigger apoptosis and modulate cell cycle. Non-conservative mutations with a potential impact in apoptosis, cell cycle arrest, caveoline-1 binding, phosphorylation, glycosylation and RNA binding were verified.
Narrative
revert the phenotype (Castaño-Rodriguez et al. 2018).
Regarding pathogenic effects, ORF3a can trigger inflammatory responses by inducing the expression of NB-κB and IL-8 (Kanzawa et al. 2006). Besides, ORF3a has been shown to be involved in inflammatory responses by promoting activation of NLRP3 inflammasome (Siu et al. 2019). NLRP3 activation is achieved by the ubiquitination of apoptosis-associated speck-like protein containing a caspase recruitment domain (ASC) facilitated by TNF receptor-associated factor 3 (TRAF3) and ORF3a direct interaction (Siu et al. 2019). Furthermore, ORF3a triggers unfolded protein response (UPR) in the endoplasmic reticulum via PKR-like ER kinase (PERK) pathway, thereby increasing IFN alpha-receptor subunit 1 (IFNAR1) phosphorylation, ubiquitination and lysosomal degradation (Minakshi et al. 2009). Thus, ORF3a may weaken INF responses and innate immunity responses. ORF3a has apoptotic and cell cycle regulation functions via caspase pathways and by arresting cells at G1 stage (Padhan et al. 2008; Yuan et al. 2007). Further, if either tyrosine (YXXφ), di-acidic (EXD), or cysteine rich motifs are mutated the apoptotic effects of ORF3a are reduced (Chan et al. 2009). Furthermore, ORF3a potassium binding activity is blocked, apoptosis is also blocked (Chan et al. 2009). From a clinical perspective, some patients recovered from SARS showed joint pain and bone damage. Consequently, the study conducted by Obitsu et al. (2009) showed that ORF3a is able to induce osteoclastogenesis and thus bone abnormalities observed in SARS survivors. Further, some patients developed pulmonary thrombosis during SARS-CoV-1 infection(Ng 2005), while ORF3a was reported to increase expression of all subunits of fibrinogen (Tan et al. 2005). Taken together, ORF3a may contribute to the formation of blood clots in some patients.
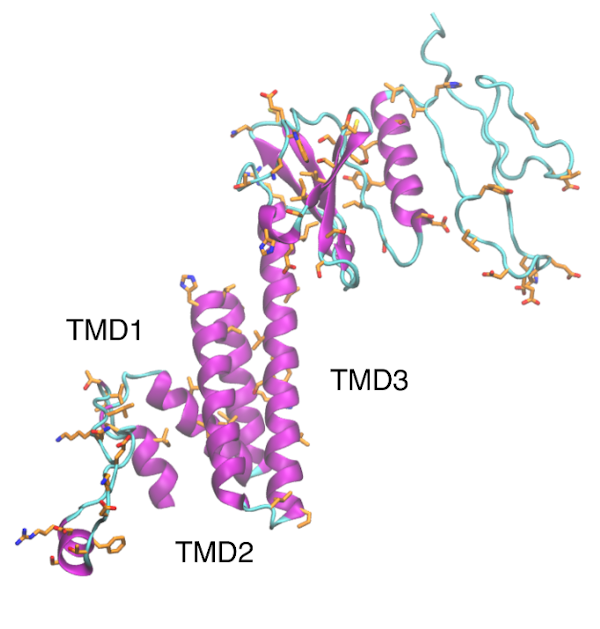
Structural analysis and comparison with SARS-CoV-1 ORF3a – ORF3a protein is the biggest of all SARS-CoV-1 accessory proteins with 274 a.a. Structurally, ORF3a possess a signal peptide (1 – 15) and three transmembrane domains TMD1 (34 – 56), TMD2 (77 – 99), and TMD3 (103 – 125) and a cytoplasmic domain at its C-terminal of 149 residues (Minakshi et al. 2014; Rota et al. 2003; R. Zeng et al. 2004). Several important residues have been identified for ORF3a function. At ORF3a cytoplasmic domain, there is a predicted calcium binding domain at residues 209 – 264 that goes through conformational changes upon binding calcium in vitro (Minakshi et al. 2014). Besides, a cysteine rich motif (Cys-Trp-Leu-Cys-Trp-Lys-Cys) from residues 127 – 133, located at the end of TMD3 region is essential for ORF3a and S protein interaction (R. Zeng et al. 2004). The tyrosine motif Tyr-Asn-Ser-Val (160 – 163) was found essential for a proper ORF3a export to the cell surface (Minakshi and Padhan 2014). Mutations at Tyr-Asn-Ser-Val motif, increase ORF3a retention at Golgi complex and lysosome degradation (Minakshi and Padhan 2014). Further, the motifs Tyr-Asn-Ser-Val, di-acidic (Glu-Gly-Asp, 172 – 173) and cysteine rich motifs, have been associated with apoptotic function of ORF3a (Chan et al. 2009). Mutations at 176 – 274 residues pointed this region to be important to induce cell cycle arrest at G1 stage (Yuan et al. 2007) Regarding subcellular transport, yeast two-hybrid assays showed that caveoline-1 interacts directly to ORF3a N-terminal domain (Padhan et al. 2007). ORF3a contains three caveolin binding sites (Trp-Gln-Leu-Ala-Leu-Tyr-Lys-Gly-Phe, Tyr-Leu-Tyr-Ala-Leu-Ile-Tyr-Phe, and Tyr-Asp-Ala-Asn-Tyr-Phe-Val-Cys-Trp) (Padhan et al. 2007) if one or two are mutated ORF3a colocalize still with caveolin-1, suggesting that only one site is enough for this interaction to take place (Padhan et al. 2007). Like M protein, ORF3a is glycosylated and substitutions Ser27Gly, Thr28Ala, Thr32Ala and Thr34Ala prevents its glycosylation (Oostra et al. 2006). Further substitutions at TMD2 Tyr91Ala and His93Ala, and TMD3 Tyr109Ala eliminated ORF3a channel activity (Castaño-Rodriguez et al. 2018). Besides, residues 125 – 200 are predicted to be β-sheet and are able to bind at the 5’-UTR the viral genomic RNA, which may in turn help with virion assembly (Sharma et al. 2007). Finally, ORF3a contains a PDZ domain-binding motif (PBM, Ser-Val-Pro-Leu, 271 – 274) required for maximum viral yield (Castaño-Rodriguez et al. 2018).
SARS-CoV-2 ORF3a is a protein of 275 residues with three transmembrane domains (TMD) and a C-terminal cytoplasmic domain. Equal to SARS-CoV-1, the predicted transmembrane positions for SARS-CoV-2 ORF3a are located at 34 – 56 (TMD1), 77 – 99 (TMD2) and 103 – 125 (TMD3). In total 76 mutations created a sequence identity of 72·36% between SARS-CoV-1 and SARS-CoV-2. The mutations are distributed along the whole ORF3a sequence, but some conservated zones were observed at TMD1, TMD2, TMD3, cysteine-rich domain and the last portion of C-terminal domain (244 – 275). TMD2 Tyr91 and His93 as well as TMD3 Tyr109 were conserved, reinforcing the idea that these residues are important for ORF3a ion channel activity. One mutation was observed in the cysteine rich motif Cys127Leu (Leu-Trp-Leu-Cys-Trp-Lys-Cys). This substitution may affect ORF3a and S protein interaction, but it is likely that the other two cysteines are enough to establish the interaction. The tyrosine motif Tyr-Asn-Ser-Val was fully conserved likely due to its importance for ORF3a function. Besides, the Glu171Ser was found in the di-acidic domain (Ser-Gly-Asp). This mutation may have a reductive effect in ORF3a apoptotic feature (Chan et al. 2009). Truncation of residues 176 – 274 impacts the role of ORF3a in cell cycle arrest.(Yuan et al. 2007) Ten mutations were detected at this zone in SARS-CoV-2 ORF3a, but also a conserved motif Asp-Tyr-Gln-Ile-Gly-Gly-Tyr-Thr-Glu with only Ser190Thr substitution was observed. Thus, this conserved motif might be enough to carry out cell cycle arrest, but this hypothesis requires proper testing.
The three caveoline-1 binding sites were highly similar (Trp-Gln-Leu-Ala-Leu-Ser-Lys-Gly-Val, Tyr-Leu-Tyr-Ala-Leu-Val-Tyr-Phe and Tyr-Asp-Ala-Asn-Tyr-Phe-Leu-Cys-Trp) with only one non-conservative substitution Tyr74Ser and two conservative, Ile112Val and Val147Leu. The Tyr74Ser substitution led us to hypothesize that residue could be a potential phosphorylation site. Although in silico analysis of phosphorylation yielded 29 putative sites, phosphorylation of ORF3a has not been demonstrated yet. Non-conservative substitutions were found in two out of four putative glycosylation sites (Ser27Asp and Thr28Phe), meanwhile Thr32 and Thr34 were conserved. This observation suggests that glycosylation of ORF3a may occur in these residues. Furthermore, Thr32 and Thr34 are found in a conserved motif (SARS-CoV-1 Val-His-Ala-Thr-Ala-Thr-Ile-Pro and SARS-CoV-2 Val-Arg-Ala-Thr-Ala-Thr-Ile-Pro) where only one conserved substitution Arg30His was found.
The ORF3a area from 125 to 200 is required for binding to the 5’-UTR of viral genomic RNA (Sharma et al. 2007). Although residues 125 – 200 is a well conserved region, several mutations were observed in this area. To address the question if any of these mutations may have an effect on ORF3a RNA binding, we paid attention to know nucleic acid binding residues arginine, lysine and histidine. Thus, one conservative Lys134Arg and non-conservative His152Asn, Lys179Ile, Lys181Glu, Arg193Trp and His194Glu were found. However, close to these mutations were found also others in their proximity that could replace their function. For instance, close to His152Asn there is a conserved histidine at position 150, next to Lys179Ile and Lys181Glu there is Glu182His, and next to Arg193Trp and His194Glu a Asp192Lys substitution was observed. These new substitutions suggest that ORF3a could retain its RNA binding properties. However this hypothesis requires empirical validation. Finally, the PBM was also found in SARS-CoV-2 ORF3a with identical sequence to SARS-CoV-1.